
連載
炭素・窒素循環におけるキープロセス~再生生産と新生産の概念の再構築~

我々人類は、様々な形で海からの恵みを享受しています。中でも、水産資源の供給、大気中の二酸化炭素濃度の調節など、人類の生存にとって不可欠な恵みの多くは、海洋内部で駆動している物資循環の絶妙なバランスの中で機能しています。その基盤を担っているのが、植物プランクトンの光合成による基礎生産です。基礎生産によって二酸化炭素と栄養塩から生み出される有機物は、食物連鎖を通じて様々な生物の成長の源となり、海洋生態系を構築し、また同時に、大気中の二酸化炭素の大きな吸収源の一つにもなっています。
一般に純一次生産(呼吸で即消費される有機物を除いた、純粋な有機物生産)量で表現される基礎生産は、使われる栄養塩の供給源の違いによって、再生生産と新生産に区別されます。再生生産とは、光合成が行われる海洋表層の有光層内部で循環する栄養塩類を用いて行われる生産、つまり、生産された有機物が有光層内で消費され、その際に再生される栄養塩類が再度基礎生産に利用されるという繰り返しによって駆動している生産、いわば“地産地消”型の生産です。再生生産は、まさに自転車操業で常にフル回転しています。そして、作り出された生産物を外に持ち出す(≒売る)ことはできませんが、その代わりに栄養塩(材料)の調達コストはかかりません。一方、有光層の系外から栄養塩類を調達して行われる生産を新生産と呼び、有光層下に沈降したプランクトンの死骸や糞などの有機物粒子が分解して生じた栄養塩類が、渦拡散、あるいは水塊の湧昇、鉛直混合等に伴って有光層内に輸送され基礎生産に使われる場合や、大気中の窒素ガスを栄養塩に変換して生産に利用する窒素固定、大気降下物による供給などが含まれます。こちらは“他産他消”型生産と言えるでしょう。有光層内において生物量や物質量がある一定期間中不変と仮定すると(いわゆる定常状態)、その間に新生産を通じて生産された有機物量は有光層の系外に輸送される事になるため、新生産は移出生産とも呼ばれます。新生産は生産物を外に持ち出す(≒売る)ことができる代わりに、栄養塩(材料)の調達コストがかかるしくみになっているわけです。
このように、海洋で営まれている基礎生産には、全く異なる栄養塩供給メカニズムを基盤とする、二つの生産が存在し、それぞれ物質循環に果たす役割が異なることが知られてきました。近年、人間活動に伴い大気中に放出された二酸化炭素ガスに対し、海洋へネットで吸収される分がどの程度あるのか、その吸収メカニズムと吸収量の見積もりが、地球温暖化の正確な将来予測のために不可欠になっています。当然、海洋表層の基礎生産は重要な吸収メカニズムの一つですが、その中で、有光層の系外、つまり大気から隔離された海洋内部へ炭素を輸送する新生産は、大気中の二酸化炭素のネットの吸収プロセスになるため“生物ポンプ”という名の下、活発に研究が進められてきました。一方、華々しい役割を演じる新生産に比べ、再生生産はやや地味な印象がありますが、亜熱帯海域のような、栄養塩を外から調達するのが非常に厳しい貧栄養な環境下においては、基礎生産を安定的に持続させ、その海域の生態系、そして生物多様性を維持するのに重要な働きをしているものと考えられます。
では、実際に再生生産と新生産はどのように区別しているのでしょうか。一般に、一次生産量は二酸化炭素(無機炭素)の固定量として測定されますが、当然これだけでは再生生産と新生産を区別することはできません。そこで、再生生産と新生産では、用いる栄養塩類の種類が理論的に異なることを利用した区別が使われてきました。すなわち、有機物内に含まれる窒素は、分解によって栄養塩類に再生される場合、まずはアンモニウム塩として再生され、有光層内では、そのまま植物プランクトン(一部細菌群集)によって再び生産に利用されます。一方、有光層内での消費を免れ有光層下深部まで輸送された有機物が、やがて分解によってアンモニウム塩に再生されると、それは硝化細菌によって最終的に化学的に安定な硝酸塩の形に変換されます。従って、このような深部の水塊が湧昇、混合などによって有光層内に貫入する場合は、植物プランクトンは硝酸塩の形で窒素を有機物生産に利用することになります。つまり、アンモニウム塩を利用する生産が再生生産で、硝酸塩を利用する生産が新生産という区別によって、基礎生産の中身を分けて評価する手法が使われてきました。
ところが、近年の研究により、新生産量がセディメントトラップ実験によって見積もられる下方への移出生産量を上回るという報告が多くされるようになり(Kawakami & Honda, 2007など)、先述した移出生産=新生産という前提が崩れ、硝酸塩の取り込みは新生産を過大評価している可能性が指摘され出しました。その要因の一つとして、有光層下部を含む、表層内での硝化の重要性が指摘されています(Yoo et al., 2006など)。硝化は光阻害の影響を受けるために、有光層内で再生されたアンモニウム塩は硝酸に変化することなく、そのまま植物プランクトンに取り込まれると考えられ、これが再生生産の定義となっていました。ところが、実際には表層に存在する硝酸塩の中に、有光層下部を含む表層内の硝化によって生成されているものが存在する事が明らかとなり、硝酸塩の取り込みによる有機物生産の中に再生生産に相当する分が含まれている可能性が指摘されています。また、トレーサー実験により植物プランクトンに取り込まれた硝酸塩の多くが溶存有機態窒素として排泄されることも報告されており(Ward & Bronk, 2001など)、これが有光層内で速やかに分解するとなれば、硝酸塩は新生産ではなく再生生産に利用されていることになります。
その一方で、有光層内への硝酸塩供給のみで評価される新生産が過小評価であるという指摘も、特に地球化学的な物質収支に基づく研究結果との比較からされています。その要因の一つとして近年注目されるようになったのが、窒素固定の再評価です(Montoya et al., 2004など) 。従来、渦拡散や湧昇、混合等による硝酸塩の供給による比べると、窒素固定の大きさは十分小さいとされていましたが、15Nトレーサーを用いた簡便で高精度な方法による報告例が増えるにつれ、特に貧栄養な低緯度海域を中心に新生産への寄与は無視できないとの結論に方向転換しつつあります。しかしながら、窒素固定のデータは海域や季節による変化が大きく(Shiozaki et al., 2009など)、今後さらに研究事例を増やす必要があります。
移出生産=新生産という概念が徐々に揺らぎつつある中、粒状有機物の鉛直輸送だけを移出生産とした場合、それが過小評価につながる事を示す研究例も近年増えてきました。特に、従来の再生生産/新生産の概念の枠組みの中では、ほとんど評価がされてこなかった溶存有機物に関しては、近年の研究の著しい進展に伴い、その概念を大きく変える可能性を秘めています。移出生産(=新生産)は、基本的に粒状有機物の鉛直輸送を想定した概念でしたが、水塊の成層期に海洋表層内に蓄積した生物生産由来の溶存有機物が、混合期になると水塊とともに鉛直輸送されるというメカニズムが、1990年代に提唱され(Carlson et al., 1994)、移出生産に溶存有機物を考慮する必要が広く認識され出しています。また、このような生物生産由来の溶存有機物の一部は、海洋の細菌群集の作用によって、短期間では分解しない(つまり二酸化炭素と栄養塩に容易に戻らない)難分解な有機物に変換されるプロセスが実験的に明らかにされ(Ogawa et al., 2001)、二酸化炭素の吸収プロセスに関わる新たな概念として“微生物炭素ポンプ”と命名されています(Jiao et al., 2010)。時間スケールの取り方に注意が必要ですが、これまで見過ごされていた新生産の一つともいえます。さらに、もう一つの見過ごされていた新生産として、動物プランクトンの鉛直移動に伴う炭素のフラックスの評価が行われ、セディメントトラップによって得られる沈降粒子のフラックスに匹敵することも報告されています(Kobari et al., 2003)。これまで、生物の死骸や糞、あるいは溶存物を主体として理解されてきた移出生産に、生きた生物自身の移動が大きく寄与する事を示す新たな概念ともいえます。
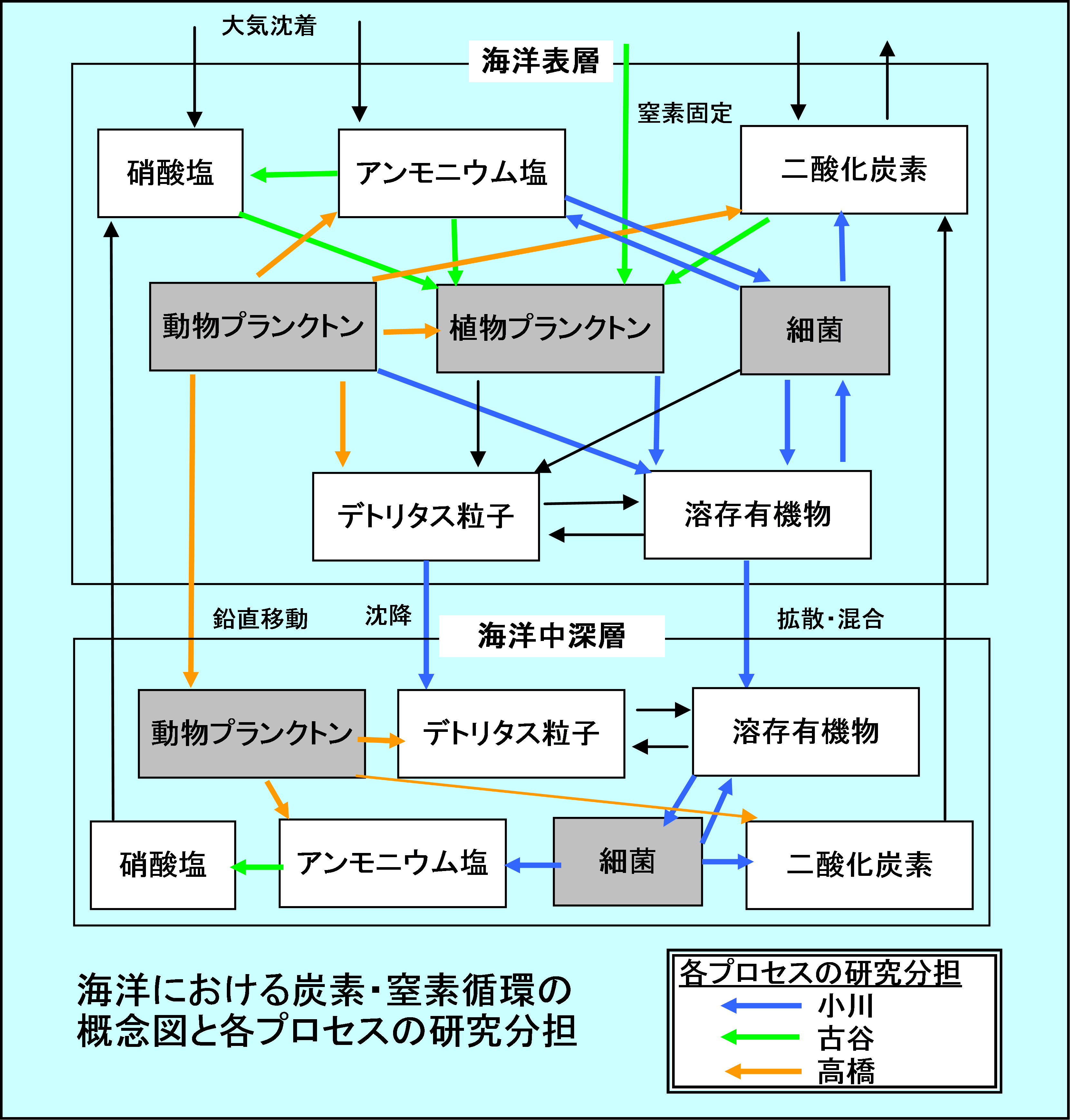
以上述べてきましたように、再生生産と新生産の従来の概念の枠組みが、近年の様々な研究成果、特に炭素・窒素循環に関わる生物地球化学的なプロセスの新発見や再考によって大きく見直されようとしています。私たちの計画研究班では、このような再生生産と新生産の概念の再構築に深く関わる素過程を、炭素・窒素循環のキープロセスと位置づけ、研究を進めています。班の構成は、研究代表の小川浩史(東大・大気海洋研究所)と、分担者が古谷 研(東大・農学生命科学研究科)および高橋一生(東大・農学生命科学研究科)の計3名からなり、それぞれの専門分野から、小川が溶存有機物と細菌群集の連鎖系、古谷が植物プランクトンを中心とした窒素循環、高橋が動物プランクトンに関わるプロセスを担当しています。図に示しましたように、海洋における炭素・窒素循環には様々なプロセスが存在し、多くの場合、各々は独立ではなく相互に関与し合って全体のバランスが保たれています。図中に各プロセスに対する担当を色分けして示していますが、この中でも特にキープロセスと位置づけ重点的に研究を進めているのが、溶存有機物⇔細菌群集の相互作用(小川)、窒素固定と植物プランクトンによる硝酸塩取り込(古谷)、動物プランクトンンによる栄養塩再生(高橋)に相当するプロセスです。私たちは、主として学術研究船白鳳丸による研究航海を利用し、太平洋における様々な海域ごとに、各プロセスに対する炭素・窒素フラックスの大きさやそれを担っている生物種を明らかにし、太平洋の“基本台帳”の情報として盛り込むことを第一の目的としています。さらに、これらの情報を相互に比較し解析することにより、各海域における、再生生産と新生産のバランスに基づく基礎生産構造の評価を行っていきます。
引用文献
Carlson, C. A., H. W. Ducklow, and A. F. Michaels (1994): Nature, 371, 405-408.
Jiao, N., G. J. Herndl, D. A. Hansell, R. Benner, G. Kattner, S. W. Wihelm, D. L. Kirchman, M. G. Weinbauer, T. Luo, F. Chen, and F. Azam (2010): Nat. Rev. Microbiol., 8, 593-599.
Kawakami, H. and M. C. Honda (2007): Deep-Sea Res. I, 54, 1070-1090.
Kobari, T., A. Shimada, and A. Tsuda (2003): Prog. Oceanogr., 57, 279-298.
Montoya, J. P., C. M. Holl, J. P. Zehr, A. Hansen, T. A. Villareal, and D. G. Capone (2004): Nature, 430, 1027-1031
Ogawa, H., Y. Amagai, I. Koike, K. Kaiser, and R. Benner (2001): Science, 292, 917-920.
Shiozaki, T., K. Furuya, T. Kodama, and S. Takeda (2009): Mar. Ecol. Progr. Ser., 377, 19-32.
Yool, A., A. P. Martin, C. Fernandez, and D. R. Clark (2007): Nature, 447, 999-1002.
Ward, B. B. and D. A. Bronk (2001): Mar. Ecol. Progr. Ser., 219, 11-24.